
A Comparative Study of Climatology, Energy and Mass Exchange in Two Forests on Contrasting Habitats in Central Siberia: Permafrost Larix gmelinii vs. Permafrost-Free Pinus sylvestris
 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Site Description
2.2. Data Processing for Net Ecosystem Exchange of CO2
3. Results
3.1. Site General and Specific Climatology
3.2. Net Radiation, Energy Balance, Bowen Ratio and Water Balance
3.3. Daily, Seasonal, and Annual Dynamics of Ecosystem CO2 Exchange: NEE, Reco, and GPP
3.4. NEE, Heat and Water Vapor Fluxes
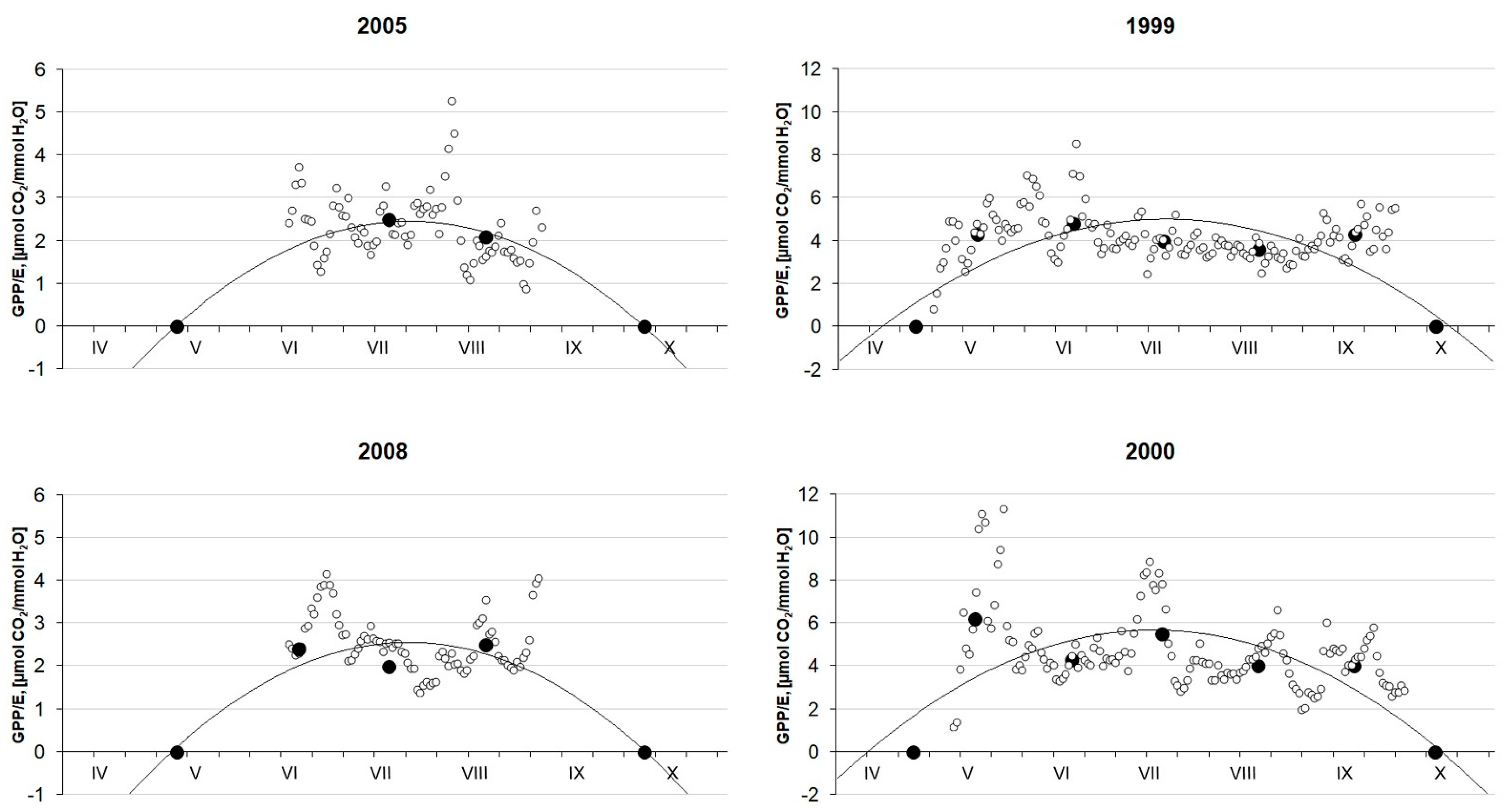
3.5. Water Use Efficiency
4. Discussion
5. Conclusions
- Two different forest ecosystems growing in contrasting habitats in interior Siberia were studied: a Pinus sylvestris forest growing on warm sandy soils and a Larix gemilii forest growing on permafrost soils with a shallow active layer depth. These forest ecosystems differ distinctively in their structure (age, height and diameter, LAI, stem density, etc.). The permafrost plays a double role: on the one hand, it supports the forest existence in a dry climate over East Siberia, delivering additional water from thawing permafrost; and on the other hand, much available energy, up to 30%–50%, is consumed in thawing ice. Thus less energy remains for sensible heat and latent heat flux, warming the soil and ambient air and for physiological processes in ecosystems.
- Net radiation was 2–2.5 fold greater in the pine forest than in the larch forest due to a 2.5 week longer growing season. Sensible and latent heat partitioned from Rn and expressed by the Bowen ratio showed that β remained at 1–2 for the growing season when the pine forest was physiologically active and increased up to 8–10 when it was not.
- Precipitation and evaporation in the pine forest were 30%–50% greater than in the larch forest. In both ecosystems, the water balance was positive for the growing season; however, the monthly and cumulative daily water balances were often negative.
- Daily maximal half-hourly CO2 fluxes were about the same in both ecosystems ~−10 µmol m–2 s–1. However, averaged daily CO2 fluxes in the pine forest were three times larger than the fluxes in the larch forest, which resulted in 228 g C m−2 season–1 vs. 83 g C m−2 season–1, respectively. The NEP patterns in both ecosystems exposed a strong signal of them being a C-sink for the growing season and year-round. Both Reco and GPP were 2–3 fold lower in our Gmelin larch.
- Water use efficiency (GPP/E) of the pine ecosystem appeared to be, on average, 2 times greater: 11 vs. 6 mg CO2 g−1H2O in the larch ecosystem. Thus the water cost per unit of C-assimilation was twice greater in the permafrost larch ecosystem.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pozdnyakov, L.K. Forestry on Permafrost; Nauka Press: Novosibirsk, Russia, 1986; p. 192. (In Russian) [Google Scholar]
- Shumilova, L.V. Botanical Geography of Siberia; Tomsk University Press: Tomsk, Russia, 1962; p. 440. (In Russian) [Google Scholar]
- Tchebakova, N.M.; Parfenova, E.I.; Soja, A.J. The effects of climate, permafrost and fire on vegetation change in Siberia in a changing climate. Environ. Res. Lett. 2009, 4, 045013. [Google Scholar] [CrossRef]
- Abaimov, A.P.; Zyryanova, O.A.; Prokushkin, S.G. Long-term investigations of larch forests in cryolithic zone of Siberia: Brief history, recent results and possible changes under Global Warming. Eurasian J. For. Res. 2002, 95–106. [Google Scholar]
- Shvidenko, A.Z.; Vaganov, E.A.; Nilsson, S. Biospheric role of Russian forests at the start of the third Millennium: Carbon budget and Kyoto Protocol. Sib. J. Ecol. 2003, 6, 649–658, (In Russian with English Summary). [Google Scholar]
- Hollinger, D.Y.; Kelliher, F.M.; Schulze, E.-D.; Vygodskaya, N.N.; Varlargin, A.; Milukova, I.; Byers, J.N.; Sogatchev, A.; Hunt, J.E.; McSeveny, T.M.; et al. Initial assessment of multi-scale measurements of CO2 and H2O flux in the Siberian taiga. J. Biogeogr. 1995, 22, 425–431. [Google Scholar] [CrossRef]
- Kelliher, F.M.; Hollinger, D.Y.; Schulze, E.-D.; Vygodskaya, N.N.; Byers, J.N.; Hunt, J.E.; McSeveny, T.M.; Milukova, I.; Sogatchev, A.; Varlargin, A.; et al. Evaporation from an eastern Siberian larch forest. Agric. For. Meteorol. 1997, 85, 135–147. [Google Scholar] [CrossRef]
- Schulze, E.-D.; Lloyd, J.; Kelliher, F.M.; Wirth, C.; Rebmann, C.; Luhker, B.; Mund, M.; Knohl, A.; Milyukova, I.; Schulze, W.; et al. Productivity of forests in the Eurosiberian boreal region and their potential to act as a carbon sink—A synthesis. Glob. Change Biol. 1997, 5, 703–722. [Google Scholar] [CrossRef]
- Ohta, T.; Hiyama, T.; Tanaka, H.; Kuwada, T.; Maximov, T.C.; Ohta, T.; Fukushima, Y. Seasonal variation in the energy and water exchanges above and below a larch forest in eastern Siberia. Hydrol. Process. 2001, 15, 1459–1476. [Google Scholar] [CrossRef]
- Dolman, A.J.; Maximov, T.C.; Moors, E.J.; Maximov, A.P.; Elbers, J.A.; Kononov, A.V.; Waterloo, M.J.; van der Molen, M.K. Net ecosystem exchange of carbon dioxide and water of far eastern Siberian larch (Larix cajanderi) on permafrost. Biogeosciences 2004, 1, 133–146. [Google Scholar] [CrossRef]
- Machimura, T.; Kobayashi, Y.; Iwahana, G.; Hirano, T.; Lopez, L.; Fukuda, M. Change of carbon dioxide budget during three years after deforestation in eastern Siberian larch forest. J. Agric. Meteorol. 2005, 60, 653–656. [Google Scholar] [CrossRef]
- Maximov, T.C. Carbon Turnover in the Larch Forests of Yakutian Part of Cryolithozone. Ph.D. Thesis, V.N. Sukachev Institute of Forests SB RAS, V.N. Sukachev Institute of Forest SB RAS, Krasnoyarsk, Russia, 2007; p. 303. (In Russian). [Google Scholar]
- Nakai, Y.; Matsuura, Y.; Kajimoto, T.; Abaimov, A.P.; Yamamoto, S.; Zyryanova, O.A. Eddy covariance CO2 flux above a Gmelin larch forest on continuous permafrost of Central Siberia during a growing season. Theor. Appl. Climatol. 2008, 93, 133–147. [Google Scholar] [CrossRef]
- Ohta, T.; Maximov, T.C.; Dolman, A.J.; Nakai, T.; van der Molen, M.K.; Kononov, A.V.; Maximov, A.P.; Hiyama, T.; Iijima, Y.; Moors, E.J.; et al. Interannual variation of water balance and summer evapotranspiration in an eastern Siberian larch forest over a 7-year period (1998–2006). Agric. For. Meteorol. 2008, 148, 1941–1953. [Google Scholar] [CrossRef]
- Nakai, Y. Net ecosystem exchange of CO2 in permafrost larch ecosystem. In Permafrost Ecosystems: Siberian Larch Forests; Osawa, A., Zyryanova, O.A., Matsuura, Y., Kajimoto, T., Wein, R.W., Eds.; Ecological Studies; Springer: Dordrecht, The Netherlands, 2010; Volume 209, pp. 183–203. [Google Scholar] [CrossRef]
- Zyryanov, V.I.; Nakai, Y.; Vaganov, E.A. Microclimate influence over CO2 fluxes in a larch stand in Central Evenkia. Conifers of the Boreal Area, 2008, V. XXV, 1–2. Available online: http://forest-culture.narod.ru/eng/journal/e_2008_1-2/zyryanov23.html (accessed on 6 February 2023).
- Olchev, A.V.; Zyryanov, V.I.; Satosina, E.M.; Fokeev, E.V.; Mukhartova, I.V.; Novenko, E.Y.; and Prokushkin, A.S. Seasonal variability of carbon dioxide, sensible and latent heat fluxes in a northern taiga larch forest of Central Siberia from eddy covariance flux measurements. Russ. Meteorol. Hydrol. 2022, 47, 804–811. [Google Scholar] [CrossRef]
- Masyagina, O.V.; Menyailo, O.V.; Prokushkin, A.S.; Matvienko, A.I.; Makhnykina, A.V.; Evgrafova, S.Y.; Mori, S.; Koike, T.; Prokushkin, S.G. Soil respiration in larch and pine ecosystems of the Krasnoyarsk region (Russian Federation): A latitudinal comparative study. Arab. J. Geosci. 2020, 13, 954. [Google Scholar] [CrossRef]
- Urban, J.; Rubtsov, A.V.; Shashkin, A.V.; Benkova, V.E. Growth, transpiration and water use efficiency of Larix sibirica, Larix gmelinii and Pinus sylvestris forest in Siberia. Acta Hortic. 2018, 1222, 124–132. [Google Scholar] [CrossRef]
- Urban, J.; Rubtsov, A.V.; Urban, A.V.; Shashkin, A.V.; Benkova, V.E. Canopy transpiration of a Larix sibirica and Pinus sylvestris forest in Central Siberia. Agric. For. Meteorol. 2019, 271, 64–72. [Google Scholar] [CrossRef]
- Virkkala, A.M.; Natali, S.; Rogers, B.M.; Watts, J.D.; Savage, J.; Connon, S.; Mauritz-tozer, M.E.; Schuur, E.A.G.; Peter, D.L.; Minions, C.; et al. The ABCflux Database: Arctic-Boreal CO2 Flux and Site Environmental Data, 1989–2020; ORNL DAAC: Oak Ridge, TN, USA, 2021. [Google Scholar] [CrossRef]
- Virkkala, A.M.; Natali, S.M.; Rogers, B.M.; Watts, J.D.; Savage, K.; Connon, S.J.; Mauritz, M.; Schuur, E.A.G.; Peter, D.; Minions, C.; et al. The ABCflux database: Arctic–boreal CO2 flux observations and ancillary information aggregated to monthly time steps across terrestrial ecosystems. Earth Syst. Sci. Data 2022, 14, 179–208. [Google Scholar] [CrossRef]
- Ueyama, M.; Iwata, H.; Harazono, Y.; Euskirchen, E.S.; Oechel, W.C.; Zona, D. Growing season and spatial variations of carbon fluxes of Arctic and boreal ecosystems in Alaska (USA). Ecol. Appl. 2013, 23, 1798–1816. [Google Scholar] [CrossRef]
- Baldocci, D.; Chu, H.; Reichstein, M. Inter-annual variability of net and gross ecosystem carbon fluxes: A review. Agric. For. Meteorol. 2018, 249, 520–533. [Google Scholar] [CrossRef]
- Natali, S.M.; Watts, J.D.; Rogers, B.M.; Potter, S.; Ludwig, S.M.; Selbmann, A.K.; Sullivan, P.F.; Abbott, B.W.; Arndt, K.A.; Birch, L.; et al. Large loss of CO2 in winter observed across the northern permafrost region. Nat. Clim. Change 2019, 9, 852–857. [Google Scholar] [CrossRef]
- Natali, S.; Watts, J.D.; Potter, S.; Rogers, B.M.; Ludwig, S.; Selbmann, P.; Sullivan, P.; Abbott, B.; Arndt, K.; Bloom, A.A.; et al. Synthesis of Winter in Situ Soil CO2 Flux in Pan-Arctic and Boreal Regions, 1989–2017; ORNL DAAC: Oak Ridge, TN, USA, 2019. [Google Scholar] [CrossRef]
- Watts, J.D.; Farina, M.; Kimball, J.S.; Schiferl, L.D.; Liu, Z.; Arndt, K.A.; Zona, D.; Ballantyne, A.; Euskirchen, E.S.; Parmentier, F.-J.W.; et al. Carbon uptake in Eurasian boreal forests dominates the high latitude net ecosystem carbon budget. Glob. Chang. Biol. 2023. [Google Scholar] [CrossRef]
- Abbott, B.W.; Jones, J.B.; Schuur, E.A.G.; Chapin, F.S., III; Bowden, W.B.; Bret-Harte, M.S.; Epstein, H.E.; Flannigan, M.D.; Harms, T.K.; Hollingsworth, T.N.; et al. Biomass offsets little or none of permafrost carbon release from soils, streams, and wildfire: An expert assessment. Environ. Res. Lett. 2016, 11, 034014. [Google Scholar] [CrossRef]
- Kajimoto, T.; Osawa, A.; Matsuura, Y.; Abaimov, A.P.; Zyryanova, O.A.; Kondo, K.; Tokuchi, N.; Hirobe, M. Individual-based measurement and analysis of root system development: Case studies for Larix gmelinii trees growing on the permafrost region in Siberia. J. For. Res. 2007, 12, 103–112. [Google Scholar] [CrossRef]
- Soil Survey Staff. Keys to Soil Taxonomy, 8th ed.; Pocahontas Press Inc.: Blacksburg, VA, USA, 1998; p. 600. [Google Scholar]
- Matsuura, Y.; Kajimoto, T.; Osawa, A.; Abaimov, A.P. Carbon storage in larch ecosystems in continuous permafrost region of Siberia. Phyton 2005, 45, 51–54. [Google Scholar]
- Wirth, C.; Schulze, E.-D.; Schulze, W.; von Stunzner-Karbe, D.; Ziegler, W.; Miljukova, I.M.; Sogatchev, A.; Varlagin, A.B.; Panvyorov, M.; Grigorev, S.; et al. Above-ground biomass and structure of pristine Siberian Scots pine forests as controlled by competition and fire. Oecologia 1999, 121, 66–80. [Google Scholar] [CrossRef]
- Tchebakova, N.M.; Kolle, O.; Zolotoukhine, D.; Arneth, A.; Styles, J.M.; Vygodskaya, N.N.; Schulze, E.-D.; Shibistova, O.; Lloyd, J. Inter-annual ad seasonal variations of energy and water vapour fluxes above a Pinus sylvestris forest in Siberian middle taiga. Tellus 2002, 54, 537–551. [Google Scholar] [CrossRef]
- Aubinet, M.; Vesala, T.; Papale, D. (Eds.) Eddy Covariance: A Practical Guide to Measurement and Data Analysis; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Pastorello, G.; Trotta, C.; Canfora, E.; Chu, H.; Christianson, D.; Cheah, Y.-W.; Poindexter, C.; Chen, J.; Elbashandy, A.; Humphrey, M.; et al. The FLUXNET2015 Dataset and the ONEFlux Processing Pipeline for Eddy Covariance Data. Sci. Data 2020, 7, 225. [Google Scholar] [CrossRef]
- Hollinger, D.Y.; Kelliher, F.M.; Byers, J.N.; Hunt, J.E.; McSeveny, T.M.; Weir, P.L. Carbon dioxide exchange between an undisturbed old-growth temperate forest and the atmosphere. Ecology 1994, 75, 134–150. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Hicks, B.B.; Meyers, T.P. Measuring biosphere-atmosphere exchanges of biologically related gases with micrometeorological methods. Ecology 1988, 69, 1331–1340. [Google Scholar] [CrossRef]
- Anthoni, P.M.; Law, B.E.; Unsworth, M.H. Carbon and water vapor exchange of an open-canopied ponderosa pine ecosystem. Agric. For. Meteorol. 1999, 95, 151–168. [Google Scholar] [CrossRef]
- Schmid, H.P.; Grimmoud, C.S.B.; Cropley, F.; Offerle, B.; Hong-Bing, S. Measurement of CO2 and energy fluxes over a mixed hardwood forest in the mid-western United States. Agric. For. Meteorol. 2000, 103, 357–374. [Google Scholar] [CrossRef]
- Wilson, K.; Goldstein, A.; Falge, E.; Aubinet, M.; Baldocchi, D.; Berbigier, P.; Bernhofer, C.; Ceulemans, R.; Dolman, H.; Field, C.; et al. Energy balance closure at FLUXNET sites. Agric. For. Meteorol. 2002, 113, 223–243. [Google Scholar] [CrossRef]
- Kukharets, V.P.; Tsvang, L.P.; Yaglom, A.M. Relationship between characteristics of turbulent surface and boundary layers of atmosphere. In Atmosphere Physics and Problems of Climate; Nauka: Moscow, Russia, 1980; pp. 162–193. [Google Scholar]
- Panin, G.N.; Bernhover, C. Parametrization of turbulent fluxes over inhomogeneous landscapes. Izv. Atmos. Ocean. Phys. 2008, 44, 755–772. [Google Scholar] [CrossRef]
- Lloyd, J.; Taylor, J.A. On temperature-dependence of soil respiration. Funct. Ecol. 1994, 8, 315–323. [Google Scholar] [CrossRef]
- Landsberg, J.J. Physiological Ecology of Forest Production; Academic Press: London, UK; Harcourt Brace Jovanovich Publishers: Orlando, NY, USA, 1986; p. 198. [Google Scholar]
- Reference Books on Climate of the USSR; Part I. Solar radiation, radiation balance and sunshine. 1966–1967; Part II. Air and soil temperature. 1966–1970; Part IV. Air humidity, precipitation and snow pack; Hydrometeoizdat: Leningrad, Russia, 1970; 20–24, Available online: http://meteo.ru/english/climate/cl_data.php (accessed on 6 February 2023).
- Tchebakova, N.M.; Parfenova, E.I.; Soja, A.J. Climate change and climate-induced hot spots in forest shifts in central Siberia from observed data. Reg. Environ. Chang. 2011, 11, 817–827. [Google Scholar] [CrossRef]
- Shibistova, O.; Zrazhevskaya, G.; Arneth, A.; Kolle, O.; Knohl, A.; Astrakhantseva, N.; Shijneva, I.; Schmerler, J. Annual ecosystem respiration budget for a Pinus sylvestris stand in central Siberia. Tellus 2002, 54, 568–589. [Google Scholar] [CrossRef]
- Morishita, T.; Masyagina, O.V.; Koike, T.; Matsuura, Y. Soil respiration in larch forests. In Permafrost Ecosystems: Siberian Larch Forests; Osawa, A., Zyryanova, O.A., Matsuura, Y., Kajimoto, T., Wein, R.W., Eds.; Ecological Studies; Springer: Dordrecht, The Netherlands, 2010; Volume 209, pp. 165–182. [Google Scholar] [CrossRef]
- Rauner, Y.L. Heat-Energy Balance of the Vegetation Cover; Hydrometeoizdat: Leningrad, Russian, 1972; p. 210. (In Russian) [Google Scholar]
- Pavlov, A.V. Energy Exchange in the Landscape Sphere of the Earth; Nauka Press: Novosibirsk, Russia, 1984; p. 256. (In Russian) [Google Scholar]
- Röser, C.; Montagnani, L.; Schulze, E.-D.; Mollicone, D.; Kolle, O.; Meroni, M.; Papale, D.; Marchesini, L.B.; Federici, S.; Valentini, R. Net CO2 exchange rates in three different successional stages of the “Dark Taiga” of central Siberia. Tellus 2002, 54, 642–654. [Google Scholar] [CrossRef]
- Ohta, T. Hydrological aspects in a Siberian larch forest. In Permafrost Ecosystems: Siberian Larch Forests; Osawa, A., Zyryanova, O.A., Matsuura, Y., Kajimoto, T., Wein, R.W., Eds.; Ecological Studies; Springer: Dordrecht, The Netherlands, 2010; Volume 209, pp. 245–269. [Google Scholar] [CrossRef]
- Hollinger, D.Y.; Kelliher, F.M.; Schulze, E.-D.; Bauer, G.; Arneth, A.; Byers, J.N.; Hunt, J.E.; McSeveny, T.M.; Kobak, K.I.; Milukova, I.; et al. Forest-atmosphere carbon dioxide exchange in eastern Siberia. Agric. For. Meteorol. 1998, 90, 291–306. [Google Scholar] [CrossRef]
- Monserud, R.A.; Onuchin, A.A.; Tchebakova, N.M. Needle, crown, stem, and root phytomass of Pinus sylvestris stands in Russia. For. Ecol. Manag. 1996, 82, 59–67. [Google Scholar] [CrossRef]
- Valentini, R.; Matteuci, A.; Dolman, A.; Schulze, E.-D.; Rebmann, C.; Moors, E.; Grander, A. Respiration as the main determinant of carbon balance in European forests. Nature 2000, 404, 861–865. [Google Scholar] [CrossRef]
- Miyazaki, S.; Ishikawa, M.; Baatarbileg, N.; Damdinsuren, S.; Ariuntuya, N.; Jambaljav, Y. Interannual and seasonal variations in energy and carbon exchanges over the larch forests on the permafrost in northeastern Mongolia. Polar Sci. 2014, 8, 166–182. [Google Scholar] [CrossRef]
- Kononov, A.V. CO2 effluxes from the permafrost soils of the larch forests depending on hydro-thermal conditions in central Yakutia. Ph.D. Thesis, Yakutsk State University, Yakutsk, Russia, 2006; p. 146. (In Russian). [Google Scholar]
- Lloyd, J.; Shibistova, O.B.; Zolotoukhine, D.; Kolle, O.; Arneth, A.; Wirth, C.; Styles, J.M.; Tchebakova, N.M.; Schulze, E.-D. Seasonal and annual variations in the photosynthetic productivity and carbon balance of a central Siberian pine forest. Tellus 2002, 54, 590–610. [Google Scholar] [CrossRef]
- Abaimov, A.P.; Lesinski, J.A.; Martinsson, O.; Milyutin, L.I. Variability and Ecology of Siberian Larch Species; Report 43; Department of Silviculture, Swedish University of Agricultural Sciences (SLU): Uppsala, Sweden, 1998; p. 123. [Google Scholar]
- Baldocchi, D.D.; Vogel, C.A.; Hall, B. Seasonal variation of carbon dioxide exchange rates above and below a boreal jack pine forest. Agric. For. Meteorol. 1997, 83, 147–170. [Google Scholar] [CrossRef]
- Jarvis, P.G.; Massheder, J.M.; Hale, S.E.; Moncrieff, J.B.; Rayment, M.; Scott, S.L. Seasonal variation of carbon dioxide, water vapor, and energy exchanges of a boreal black spruce forest. J. Geophys. Res. 1997, 120, 28953–28966. [Google Scholar] [CrossRef]
- Aurela, M.; Tuovinen, J.P.; Laurila, T. Net CO2 exchange of a subarctic mountain birch ecosystem. Theor. Appl. Climatol. 2001, 70, 135–148. [Google Scholar] [CrossRef]
- Li, S.-G.; Asanuma, J.; Kotani, A.; Eugster, W.; Davaa, G.; Oyunbaatar, D.; Sugita, M. Year-round measurements of net ecosystem CO2 flux over a montane larch forest in Mongolia. J. Geophys. Res. 2005. [Google Scholar] [CrossRef]
- Black, T.A.; den Hartog, G.; Neumann, H.H.; Blanken, P.D.; Yang, P.C.; Russel, C.; Nesic, Z.; Lee, X.; Chen, S.G.; Staebler, R.; et al. Annual cycles of water vapour and carbon dioxide fluxes in and above a boreal aspen forest. Glob. Chang. Biol. 1996, 2, 219–229. [Google Scholar] [CrossRef]
- Abaimov, A.P.; Prokushkin, S.G.; Zyryanova, O.A.; Kaverzina, L.N. Peculiarities of forming and functioning larch forests on frozen soils. Lesovedenie 1997, 5, 13–23. (in Russian). [Google Scholar]
- Vygodskaya, N.N.; Milyukova, I.; Varlagin, A.; Tatarinov, F.; Sogachev, A.; Kobak, K.I.; Desyatkin, R.; Bauer, G.; Hollinger, D.Y.; Kelliher, F.M.; et al. Leaf conductance and CO2 assimilation of Larix gmelinii under natural conditions of Eastern Siberian boreal forest. Tree Physiol. 1997, 17, 607–615. [Google Scholar] [CrossRef]
- Bryukhanova, M.V.; Kirdyanov, A.V.; Prokushkin, A.S.; Silkin, P.P. The features in the xylem genesis of Larix gmelinii under cryolithic conditions of Central Siberia. Russ. J. Ecol. 2013, 5, 323–329. (In Russian) [Google Scholar]
- Orlov, A.Y.; Koshelkov, S.P. Soil Ecology of Scots Pine; Nauka: Moscow, Russia, 1971; p. 322. (In Russian) [Google Scholar]
- Mahli, Y.; Baldocchi, D.; Jarvis, P. The carbon balance of tropical, temperate and boreal forests. Plant Cell Environ. 1999, 22, 715–740. [Google Scholar]
- Yu, G.; Song, X.; Wang, Q.; Liu, Y.; Guan, D.; Yan, J.; Sun, X.; Zhang, L.; Wen, X. Water-use efficiency of forest ecosystems in eastern China and its relations to climatic variables. New Phytol. 2007, 177, 927–937. [Google Scholar] [CrossRef]
- Morén, A.-S.; Lindroth, A.; Grelle, A. Water-use efficiency as a means of modelling net assimilation in boreal forests. Trees 2001, 15, 67–74. [Google Scholar] [CrossRef]
- Lindroth, A.; Cientiala, E. Water use efficiency of short-rotation Salix viminalis at leaf, tree and stand scales. Tree Physiol. 1996, 16, 257–262. [Google Scholar] [CrossRef]
- Tchebakova, N.M.; Parfenova, E.I. The 21st century climate change effects on the forests and primary conifers in central Siberia. Bosque 2012, 33, 253–259. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Larch Ecosystem | Pine Ecosystem | |||||
|---|---|---|---|---|---|---|
| 2005 | 2007 | 2008 | 1998 | 1999 | 2000 | |
| P | 122 | 184 | 215 | 248 | 233 | 228 |
| E | 112 | 127 | 116 | 173 | 192 | 200 |
| P–E | 10 | 57 | 99 | 75 | 41 | 28 |
| Larch Ecosystem | Pine Ecosystem | |||||
|---|---|---|---|---|---|---|
| 2005 | 2007 | 2008 | 1998 | 1999 | 2000 | |
| GPP, gCm−1 | 167 | 186 | 210 | 543 | 592 | 552 |
| Reco, gCm−1 | 104 | 106 | 104 | 324 | 341 | 340 |
| NEP, gCm−1 | 63 | 80 | 106 | 219 | 251 | 212 |
| Ecosystem | R2 (p < 0.001) | |||
|---|---|---|---|---|
| H | LE | Ro | ||
| Larch | 2004 | −0.5 | 0.8 | 0.7 |
| 2005 | −0.6 | 0.4 | 0.8 | |
| 2007 | −0.7 | 0.3 | 0.8 | |
| 2008 | −0.8 | 0.4 | 0.8 | |
| Pine | 1998 | −0.9 | 0.9 | 0.9 |
| 1999 | −0.6 | 0.7 | 0.7 | |
| 2000 | −0.6 | 0.7 | 0.7 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tchebakova, N.M.; Zyryanov, V.I.; Zyryanova, O.A.; Parfenova, E.I.; Kajimoto, T.; Matsuura, Y. A Comparative Study of Climatology, Energy and Mass Exchange in Two Forests on Contrasting Habitats in Central Siberia: Permafrost Larix gmelinii vs. Permafrost-Free Pinus sylvestris. Forests 2023, 14, 346. https://doi.org/10.3390/f14020346
Tchebakova NM, Zyryanov VI, Zyryanova OA, Parfenova EI, Kajimoto T, Matsuura Y. A Comparative Study of Climatology, Energy and Mass Exchange in Two Forests on Contrasting Habitats in Central Siberia: Permafrost Larix gmelinii vs. Permafrost-Free Pinus sylvestris. Forests. 2023; 14(2):346. https://doi.org/10.3390/f14020346
Chicago/Turabian StyleTchebakova, Nadezhda M., Viacheslav I. Zyryanov, Olga A. Zyryanova, Elena I. Parfenova, Takuya Kajimoto, and Yojiro Matsuura. 2023. "A Comparative Study of Climatology, Energy and Mass Exchange in Two Forests on Contrasting Habitats in Central Siberia: Permafrost Larix gmelinii vs. Permafrost-Free Pinus sylvestris" Forests 14, no. 2: 346. https://doi.org/10.3390/f14020346